Guide L-111
Louis C. Bender
College of Agricultural, Consumer and Environmental Sciences, New Mexico State University
Author: Senior Research Scientist (Natural Resources), Department of Extension Animal Sciences and Natural Resources, New Mexico State University. (Print Friendly PDF)
Wildlife enterprises are increasingly important for ranch income, and in many cases provide the highest net revenues for landowners. Prices for high-end elk and mule deer hunts can exceed $15,000 per animal, and even less intensive operations can easily receive more than $2,500 for mule deer. The high revenue from wildlife enterprises has heightened competition among landowners (as well as state agencies, based on proliferation of “Governor’s Tags” and similar authorizations) for the hunter’s dollar, and this competition has resulted in significant interest in increasing the quality (i.e., antler or horn size) of elk, mule deer, and other ungulates. Attempts to improve quality (quality management; QM) can include establishing habitat improvement or feeding programs, purchasing semen from “superior” sires, and even purchasing breeding stock, often at more than $50,000 per individual. While these and other practices can increase overall herd quality, there is often a trade-off between returns and costs. Additionally, while many practices may work with intensive breeding or high fence operations, they may fare poorly in traditional extensive ranching operations. Finally, a “more is better” approach meshes poorly with QM, so more intensive operations may actually produce fewer quality animals, or may produce them for neighboring areas because of increased dispersal. Hence, a basic understanding of what is and is not important and appropriate is needed for those considering a QM approach.
The keys to QM, in order of importance, are (1) age structure, (2) nutrition, and (3) genetics. Population age structure affects both productivity and development of secondary sexual characteristics such as horns, pronghorns, and antlers; to keep things simple, this guide will primarily discuss antlers, although the management concepts are similar for all species. At its most basic, males simply have to live long enough to grow large antlers. More specifically, males will not maximize antler size until after body mass has been maximized because body size, more commonly than antler size, determines reproductive success. Additionally, body mass and antler mass are positively correlated, so bigger males tend to grow bigger antlers (Figure 1).

Figure 1. Trophy bull elk? Maybe…maybe not. Note the small body size of this adult bull, which makes his antlers appear larger than they actually are because of the smaller body-associated visual references. (Photo courtesy Jason Schlie.)
Age structure
What is not often appreciated is just how old bulls and bucks must be to maximize antler development. Elk tend to peak antler development at age 8 to 10, about 2 to 3 years after they peak body mass (Figure 2). Mule deer will peak antler mass at age 5 to 7, again 2 to 3 years after peaking body mass. All will begin to lose antler configuration after the peak ages, although mass may remain high. The exception to this is pronghorn, which often peak prong development as early as age 3.
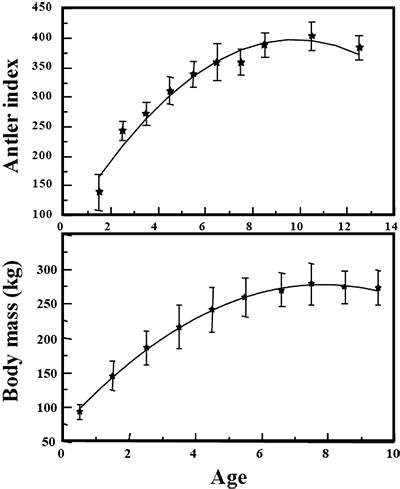
Figure 2. Common to most ungulates, the peak in antler development in elk occurs 2 to 3 years after the peak in body mass. (Data from Bender [2008].)
Quality management requires that individuals live long enough to reach these optimal ages, which is no easy task. The proportion of the population surviving to optimal ages is determined by the overall mortality rate of males. Because harvest makes up the majority of male mortality, harvest rates must therefore be low enough to allow survivorship well beyond age 5. The question is how low, and how do you achieve these lower harvest rates?
In general, annual male survival rates must be above 60% to get sufficient numbers of bucks into older age classes (Figure 3), assuming that harvest is proportional across age classes. The more selective the harvest is for older males, the lower the harvest rate must be to allow individuals to reach optimal ages. There is no panacea for getting these high survival rates other than remembering that the only way to have older males is to not kill them (Figures 3 and 4). Antler-point restrictions, or selective harvest criteria (SHC), such as 3- or 4-point minimums that protect younger males from harvest, are frequently touted to increase survival of older males. For example, many practitioners of Quality Deer Management (QDM) in the Southeast use this approach. However, it is NOT the antler restriction that results in more mature bucks in QDM; rather, it is severely limiting the numbers of adults harvested, usually to less than 30% of the age 2 and older population. This is key, because if overall harvest is NOT limited under a 3- or 4-point restriction, the result is often elimination of virtually all males older than age 3 (the only males that can be harvested with the SHC). Remember, it is the overall mortality (harvest) rate that determines survivorship—not any SHC. Any harvest strategy can—or cannot—produce trophy males, depending on just how many males are harvested (Figure 4). More discussion about an “optimal harvest strategy” will follow below.
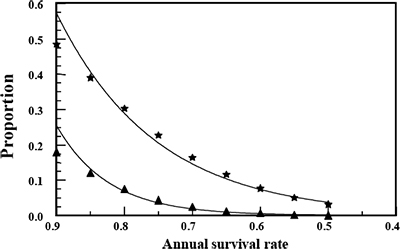
Figure 3. Proportion of males surviving to age 5 («) or age 10 (p) given various annual male survival rates assuming that harvest is proportional across age classes. Notice that less than 10% of males live to age 5 until survival reaches more than 60%, and less than 10% reach age 10 until survival reaches more than 85%. (Data from Bender [2008].)
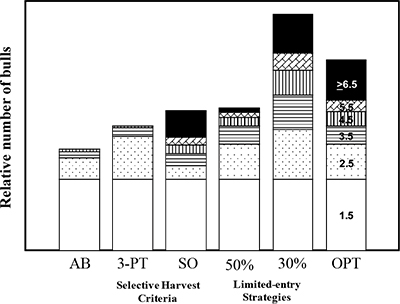
Figure 4. Preseason age structure of ungulates under common harvest strategies. Strategies include open entry-any bull (AB), two common selective harvest strategies (open-entry 3-point or better only [3-PT] and open-entry spike bull with branch antlered bulls by limited entry permit only [SO]), and two limited-entry strategies (one designed to limit overall bull mortality rate to 0.50 [50%], the other designed to limit overall bull mortality to 0.30 [30%]). Also shown is an optimal (OPT) strategy that allows high survivorship into older age classes (i.e., 6-years-old and older) but limits overall bull numbers by harvesting younger bulls at a ratio of 3–5:1, with an overall mortality rate of 0.35–0.40. Note that only strategies that limit overall mortality of bulls severely (i.e., 30%, OPT), or at least mortality of branch antlered bulls (SO), allow survivorship into trophy age classes.
How do you know what the mortality rate is? Age structure gives a good approximation. If you perform prehunt composition surveys, look for the percent of the herd that are yearlings; this will approximate the overall mortality rate (Bender, 2006). If you keep harvest data, look at the proportions between age classes in the harvest; this will do the same (Bender, 2006). Last, because the adult sex ratio (ASR) is simply the result of female mortality rate divided by male mortality rate, you can use ASR to approximate mortality rates as well. Male/female ratios of more than 60/100 in stable or increasing herds result from adequately low male mortality (see Bender, 2006 for details).
Managing for older age structure does have behavioral challenges. Lower male mortality rates result in more adult males in the population, which in turn increases stress from social interactions (and possibly less nutrition if overall population densities are not controlled). Increased stress and lower nutrition both compromise the immune system and can lower the quality of antlers produced (Ozoga and Verme, 1982). High stress caused by high adult male densities also results in more dispersal, and possibly more antler damage from frequent social dominance interactions. Dispersal is a particular concern in QM because males in trophy age classes can be physically dominated by younger but physically mature males. Deer breeders in the eastern U.S. and in Europe, where programs designed to produce “super bucks” have the longest history, particularly try to limit stress by limiting the numbers of adult males in close proximity (i.e., in the same pens, holding pastures). Because stress hormones from constant social interactions can affect size and symmetry of antlers, managing densities is critical for optimizing antlers, especially in more intensive operations.
Stress, lowered nutrition, and unintended genetic influences can also decrease body sizes, and, in general, larger males grow bigger antlers. For example, antler mass is proportional to body mass~1.6 in red deer (elk; Hyvarinen et al., 1977), and there are strong correlations (r > 0.94) among all cervids between metabolic body mass and antler length (Bubenik, 1985). Body mass is primarily an age and nutritional effect, although body mass also has moderate heritability and thus can also be influenced by genetics.
So, what is the best harvest strategy given the age structure, density, and behavioral issues discussed above? It is a strategy that does not allow males to get too numerous (to minimize dispersal and other behavioral issues) while allowing males to survive into older age classes (Figure 4). Such an “optimal harvest strategy” has higher overall mortality on males than highly restrictive strategies (such as the 30% limited-entry strategy in Figure 4), but maintains the numbers reaching trophy age classes by directing much of the harvest mortality at younger males. Thus, overall mortality rates increase, decreasing ASRs and total number of males, but age structure is maintained. Such an optimal strategy would impose an overall male mortality rate of 40% or so, but allocate harvest between younger (i.e., management) males and trophy males at a ratio of 3–5 management males harvested for each trophy. This strategy allows wildlife enterprises to maintain revenues from trophy males, increase revenues from younger males, and increase the likelihood that the trophies they produce remain on their property.
“What exactly is a ‘management’ buck or bull?” is a frequent question I receive. The short answer is, anything that is not a trophy. Aside from that, there is no simple definition. Some managers will cull out younger males with smaller antlers, under the belief that the antlers will remain small throughout life. Others will cull out atypical antler configurations, based on the same beliefs about heritability of such features. The reality is that these and other questions regarding heritability of antlers and other genetic issues are far from clear. These will be discussed later in the Genetics section of this guide. Last, some will target very old males that are regressing in their antler development, while others will treat these as trophies. So, aside from not being a trophy-aged male, a “management” bull or buck simply depends on the eye of the beholder.
Nutrition
As early as the 1940s, pioneering nutritional work by Franz Vogt in Germany demonstrated the overwhelming effect of nutrition on antler development (summarized in Geist, 1986). Vogt found that pregnant deer fed a supplement high in protein, energy, and minerals over generations produced superior stags even if the original genetic stock was considered “mediocre.” For example, one group doubled in body mass and more than doubled in antler mass in just three generations. Clearly, nutrition must be optimal for either age or genetic effects to be seen in antlers.
Unfortunately, nutritional requirements for antler development in wild herbivores are poorly understood. Some general guidelines likely apply, however.
-
As noted above, body mass will be maximized before antlers because antlers are a luxury tissue. So, while nutrition can maximize potential within an age class, it cannot compensate for survival to optimal ages.
-
Nutritional requirements for optimal antler growth probably mirror needs of females for lactation. These needs are high: for elk, diets of more than 67% digestibility and more than 16% protein, and more than 70% digestibility and more than 18% protein for deer.
-
While little firm knowledge is available on mineral requirements for antler growth, anecdotal evidence suggests that mineral supplements can enhance development.
Nutrition is best managed through proper habitat management, habitat enhancements, and animal densities; this concept is called “density dependence.” Higher densities mean fewer resources (food, space, etc.) for each individual, which in turn results in less productivity, whether productivity is defined as young produced, size of individuals, or antler development. That means overall herd size must be controlled, which is not easy for free-ranging operations in states like New Mexico where no female harvest is permitted for species such as mule deer. This is especially problematic because low mortality rates result in an older population age structure, which can limit the overall productivity of herds, and thus economic returns, because fecundity of females declines with age (Figure 5; Bender, 2008; Bender and Hoenes, 2018). Keeping female densities low can increase habitat quality for males as well as overall productivity of herds. The same is true for males; selective harvesting of younger males increases resources available for older males.
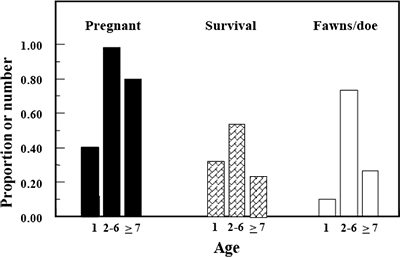
Figure 5. Pregnancy, fawn survival, and number of fawns eventually recruited per doe decline with age of doe mule deer in southern New Mexico (Bender and Hoenes, 2018). The reproductive senescence seen in older does (age 7 and older) is underestimated in this data because the sample contained few does older than age 10. Mule deer does can live more than 15 years, so the majority of does in unharvested populations show decreased reproductive potential.
In arid regions like New Mexico, even optimal habitat management may result in poor antler expression because of frequent annual and seasonal droughts. Consequently, many managers feed or supplement wildlife to maintain individual quality even during years of poor habitat productivity. To be biologically and economically effective, supplementation should provide the individual with nutrients they need when they need them. Nutrient needs, in turn, are determined by the timing, composition, and growth of antlers. During their active growth phase, antlers are approximately 80% protein, clearly indicating the need for a high-protein supplement. During mineralization and after hardening, antlers are approximately 60% mineral (still 40% protein), primarily calcium (Ca), phosphorous (P), and magnesium (Mg). To grow antlers, individuals must get these nutrients from their feed or from their body. While individuals can frequently get adequate protein from forage, they usually mobilize most minerals from their bones, particularly the ribs, reserves that are stored throughout the year.
Supplementation with a high-protein feed—at least 18%, but more than 20% is better—and a good mineral mix that includes Ca, P, and Mg, as well as vitamin D (or its precursors), can provide individuals with all their needs to maximize their antler potential. For the mineral mix, a 2:1 Ca:P ratio is probably adequate, with Mg present in lesser amounts (maybe at 2:1:0.1 to 0.3). Vitamin D or precursors potentially aid dietary mineral absorption, although the mechanisms involved in heightened mineral absorption in wild ungulates are unclear (Brown, 1990). The exact quantities in the mineral mix are likely less important than the ratios and covering the basic nutrients listed above; wild herbivores will regulate their intake to satisfy their cravings.
Late April through September is the critical time for antler growth (the March–June period is similarly critical for adult survival and production of juveniles), and is the period to supplement if desired. Note that there is no need to feed all year unless you are trying to artificially increase carrying capacity of the operation; however, this practice is NOT recommended because wild ungulates will preferentially use green feed as palatability increases, leading to overuse of natural forage. However, if feeding later or throughout the year, managers should decrease protein and increase energy as animals look to build up body fat later in the year, not muscle mass, and the metabolic use of protein for energy in ruminants is energetically inefficient. Continuing a high-protein supplement past September has little effect on males, but makes it more difficult for females to recover condition during and after weaning. It can, however, dramatically improve juvenile growth and survival. Last, remember to either introduce animals to the supplement slowly in late winter or provide it in lesser amounts throughout the year to allow digestive physiology to adjust to the supplement.
Reports of antler breakage, especially with elk, are becoming more common on many wildlife enterprises in New Mexico, and broken antlers obviously decrease the trophy value of bulls. Exactly what increases the likelihood of antler breakage is not well understood (e.g., Chapman, 1980; McDonald et al., 2005; Johnson et al., 2007), but may be related to antler density (i.e., specific gravity). Specific gravity of antlers can vary with location on the antler, age of individuals, body mass, nutrition, and population density, among other factors (Hyvarinen et al., 1977; Miller et al., 1985). Specific gravity is determined primarily by the ratio of hardened bone (the dense, compacted bone around the exterior of an antler) to trabecular bone (the porous bone in the interior of an antler) in the antler. It has been hypothesized that antlers may be less prone to breakage if specific gravity is lower (because liquid in the bony pores has a “shock absorbing” effect) or higher (because denser, compact bone has greater resistance to breakage)(Chapman, 1980), highlighting the uncertainty regarding the underlying reasons for antler breakage.
Although preliminary, data I have collected suggest that specific gravity of broken antlers from elk was lower than specific gravity of unbroken antlers. If this is the case, then breakage may be minimized with increased nutrition (Hyvarinen et al., 1977; Johnson et al., 2007) since increased nutrition is positively associated with higher specific gravity and heavier body mass (and increased body mass is positively related to increased antler mass as well). Mineral supplementation during the active growth period of antlers may also be useful to relieve any deficiencies that may contribute to lower specific gravity. Calcium in particular has consistently been shown to increase the strength of bone in a variety of mammals and is the primary mineral in hardened antler, but maintaining proper mineral ratios in supplements is critical (see above).
As noted above, the April–September period is key to antler development, and providing a quality mineral supplement freely may increase mineralization in bulls. Also, prescribed burning in spring increases mineral content of forages (as well as protein content, digestibility, and overall forage biomass) (Bender, 2011). Implementing a burning program in late winter or early spring increases minerals available to bulls through both forage as well as geophagy (consumption of ash following burning). A burning program may be the most cost-effective way to provide additional minerals to bulls.
Genetics
Things become more complicated and even more uncertain with the genetics of antler development. The strength of the link between genetics and quality can be very blurry; most work on genetics is hopelessly confounded by superior nutrition and low stress typical of studies of penned wildlife. Despite this, a couple of conclusions can be drawn. First, genetic potential can be fully expressed if and only if nutrition is superior and individuals reach optimal ages. If age structure is lacking and individuals cannot achieve large body sizes, there is much less opportunity for antler genetics to contribute to quality. Second, be aware that management for “optimal” genetics can have unintended consequences as well.
Most of the genetic work on antler development has involved white-tailed deer in penned studies at multiple public and private facilities. Results have generally shown that bucks with superior mass and antler development as yearlings maintained larger body sizes and better antler development throughout their lives, with body mass averaging 10 to 25% greater and antler mass more than 50% heavier by age 5 and older (e.g., Harmel, 1982). Progeny of these superior bucks similarly had a greater tendency to be branched as yearlings, and some subsequent genetic analyses indicated that most antler traits (as well as body mass) appear to have moderate to high heritability (Williams et al., 1994). Thus, trials that selected only the best yearlings for breeding under identical diets were able to gradually increase the average antler size of the study herds (e.g., Frels et al., 2002). Similar results have been seen with elk and red deer on private ranches, though little has been done with mule deer.
However, these results are not invariant. Work with free-ranging white-tailed deer in Texas found no relationship between antler quality as yearlings and mature deer (Koerth and Kroll, 2008). In a penned study at Texas A&M, a superior sire produced 4 spikes and 4 yearling offspring with 4 to 11 points despite identical high-quality diets (Brown, 1990). Similarly, research in Mississippi found that environment had a much greater effect on antler development of yearlings than did the sire, as well as low heritability of antler characteristics (Lukefahr and Jacobson, 1998). Work on free-ranging red deer also showed low to moderate heritability of antler mass (Kruuk et al., 2002). Further complicating this, males receive half of their genetic propensity for antlers from their mother, for which genetic potential is frequently unknown.
I have also noticed that highly selective breeding operations often produce superior antler development in younger animals, but this early advantage is lost by the time individuals reach their age of maximum antler development. These observations are partially influenced by nutritional status as well as age, because most intensive breeding programs keep individuals at high density, limiting their nutrition as well as elevating social stress. Further, many of the superior individuals get harvested at younger ages because of their antler development (e.g., shot as a 350-inch bull at age 4–5, 3 to 4 years before peak).
Along with inconsistent results on early antler expression, there is also the potential for unintended deleterious effects when breeding for superior genetics. In several cases, I have noticed that bucks and bulls from intensive breeding programs showed smaller body sizes, and, as noted above, there is a positive correlation between body mass and antler mass. Similarly, use of SHC may potentially negatively affect the genetic stock as well, although this is debated (e.g., Heffelfinger, 2017). For example, if heritability of antler characteristics is at least moderate and future antler quality can be predicted based on yearling development, SHCs selecting bucks with 3 or more antler points can potentially remove the superior bucks from a herd by harvesting yearlings that show branched antlers in areas where yearlings are able to maximize their antler potential (i.e., areas of high habitat quality, fenced operations, etc.) (Harmel, 1982; Strickland et al., 2001).
Summary
So, how to implement a QM strategy? First, QM will work best where habitat quality is high; the most important aspect of high-quality habitat is good available nutrition (nutrition effect). Second, responses will be faster as the male mortality (harvest) rate becomes lower (age effect). Third, overall population densities—both male and female—need to be kept low to (1) maximize resource capture and hence diet quality, (2) minimize competition among males and between males and females, (3) minimize stress and social encounters that may lead to antler breakage, and (4) minimize dispersal. These density effects can be mitigated by supplementation programs, although it must be emphasized that feeding will affect only resource capture, so it alone is only a partial solution. Finally, there are few—probably no—areas that lack the genetic potential in existing herds to grow trophy bulls and bucks in New Mexico. Aside from high-intensity breeding or high fence operations, resources are best invested in enhanced nutrition and age structure for QM. Even intensive operations probably could increase their quality by focusing on nutrition and stress; few studies have shown a measurable gain from adding “superior” individuals to established herds (Miller and Marchinton, 1995).
Selected literature
Bender, L.C. 2006. Uses of herd composition ratios in ungulate management. Wildlife Society Bulletin, 34, 1225–1230.
Bender, L.C. 2008. Age structure and population dynamics. In S.E. Jorgensen and B.D. Fath (Eds.), Encyclopedia of Ecology, 1st ed., vol. 1 (pp. 65–72). Amsterdam, The Netherlands: Elsevier B.V.
Bender, L.C. 2011. Burning for big game [Circular 657]. Las Cruces: New Mexico State University Cooperative Extension Service. https://pubs.nmsu.edu/_circulars/CR657/
Bender, L.C., and B.D. Hoenes. 2018. Age-related fecundity of mule deer in south-central New Mexico. Mammalia, 82, 124–132.
Brown, R.D. 1990. Nutrition and antler development. In G.A. Bubenik and A.B. Bubenik (Eds.), Horns, pronghorns, and antlers(pp. 426–441). New York: Springer-Verlag.
Bubenik, A.B. 1985. Reproductive strategies in cervids. In P.F. Fennessy and K.R. Drew (Eds.), Biology of deer production (pp. 367–374). Royal Society of New Zealand, Bulletin 22.
Bubenik, G.A., and A.B. Bubenik (Eds.). 1990. Horns, pronghorns, and antlers. New York: Springer-Verlag.
Chapman, D.I. 1980. Antler structure and function—A hypothesis. Journal of Biomechanics, 14, 195–197.
Frels, Jr., D.B., E. Fuchs, and W. Armstrong. 2002. Genetic and environmental interaction in white-tailed deer. Final Report, Federal Aid Project W-127-R-9 Job 96. Austin: Texas Department of Wildlife.
Geist, V. 1986. Super antlers and pre-World War II European research. Wildlife Society Bulletin, 14, 91–94.
Harmel, D.E. 1982. Effects of genetics on antler quality and body size in white-tailed deer. In R.D. Brown (Ed.), Antler development in Cervidae (pp. 339–348). Kingsville, TX: Caesar Kleberg Wildlife Research Institute.
Heffelfinger, J.R. 2017. Inefficiency of evolutionarily relevant selection in ungulate trophy hunting. Journal of Wildlife Management, 82, 57–66. doi: 10.1002/jwmg.21337
Hyvarinen, H., R.N.B. Kay, and W.J. Hamilton. 1977. Variation in weight, specific gravity, and composition of antlers of red deer (Cervus elaphus). British Journal of Nutrition, 38, 301–311.
Johnson, H.E., V.C. Bleich, and P.R. Krausman. 2007. Mineral deficiencies in Tule elk, Owens Valley, California. Journal of Wildlife Diseases, 43, 61–74.
Koerth, B.H., and J.C. Kroll. 2008. Juvenile-to-adult antler development in white-tailed deer in south Texas. Journal of Wildlife Management, 72, 1109–1113.
Kruuk, L.E.B., J. Slate, J.M. Pemberton, S. Brotherstone, F. Guinness, and T. Clutton-Brock. 2002. Antler size in red deer: Heritability and selection but no evolution. Evolution, 56, 1683–1695.
Lukefahr, S.D., and H.A. Jacobson. 1998. Variance component analysis and heritability of antler traits in white-tailed deer. Journal of Wildlife Management, 62, 262–268.
McDonald, C.G., S. Demarais, T.A. Campbell, H.F. Janssen, V.G. Allen, and A.M. Kelley. 2005. Physical and chemical characteristics of antlers and antler breakage in white-tailed deer. Southwestern Naturalist, 50, 356–362.
Miller, K.V., and R.L. Marchinton. 1995. Quality whitetails: The why and how of quality deer management. Mechanicsberg, PA: Stackpole Books.
Miller, K.V., R.L. Marchinton, J.B. Beckwith, and P.B. Bush. 1985. Variations in density and chemical composition of white-tailed deer antlers. Journal of Mammalogy, 66, 693–701.
Ozoga, J.J., and L.J. Verme. 1982. Physical and reproductive characteristics of a supplementally-fed white-tailed deer herd. Journal of Wildlife Management, 46, 281–301.
Strickland, B.K., S. Demarais, L.E. Castle, J.W. Lipe, W.H. Lunceford, H.A. Jacobson, D. Frels, and K.V. Miller. 2001. Effects of selective harvest strategies on white-tailed deer antler size. Wildlife Society Bulletin, 29, 509–520.
Williams, J.D., W.F. Krueger, and D.H. Harmels. 1994. Heritabilities for antler characteristics and body weights in yearling white-tailed deer. Heredity, 73, 78–83.
Acknowledgments
I thank R. Baldwin, University of California-Davis; J. Boren, New Mexico State University; A. Darrow, Mexico Coast Land and Cattle; J. Heffelfinger, Arizona Game and Fish Department; B. Hoenes, Washington Department of Fish and Wildlife; and Octavio Rosas, Colegio de Postgraduados, San Luis Potosí, Mexico, for reviewing the original or revised versions of this publication.
For further reading
CR-657: Burning for Big Game
https://pubs.nmsu.edu/_circulars/CR657/
CR-662: Guidelines for Management of Habitat for Mule Deer
https://pubs.nmsu.edu/_circulars/CR662/
CR-664: Guidelines for Monitoring Elk and Mule Deer Numbers in New Mexico
https://pubs.nmsu.edu/_circulars/CR664/
CR-669: Using Population Ratios for Managing Big Game
https://pubs.nmsu.edu/_circulars/CR669/

Lou Bender is a Research Scientist (Wildlife) with the Department of Extension Animal Sciences and Natural Resources at NMSU. He earned his Ph.D. from Michigan State University. His research and management programs emphasize ungulate and carnivore management, integrated wildlife and livestock habitat management, and wildlife enterprises in the Southwest and internationally.
To find more resources for your business, home, or family, visit the College of Agricultural, Consumer and Environmental Sciences on the World Wide Web at pubs.nmsu.edu
Contents of publications may be freely reproduced for educational purposes. All other rights reserved. For permission to use publications for other purposes, contact pubs@nmsu.edu or the authors listed on the publication.
New Mexico State University is an equal opportunity/affirmative action employer and educator. NMSU and the U.S. Department of Agriculture cooperating.
Revised April 2018 Las Cruces, NM