Circular 657
Revised by Louis C. Bender
College of Agricultural, Consumer and Environmental Sciences, New Mexico State University
Author: Senior Research Scientist (Natural Resources), Department of Extension Animal Sciences and Natural Resources, New Mexico State University. (Print Friendly PDF)
Prescribed burning is a management tool that is used to alter the composition and structure of vegetation on public and private lands in New Mexico. Burning is frequently used to increase habitat quality for big game species, such as mule deer (Odocoileus hemionus), elk (Cervus elaphus), and pronghorn (Antilocapra americana), and can be an economically viable alternative to more costly management practices, such as forage plots or feeding, for increasing the quality of wildlife habitat and economics of wildlife enterprises (Bender, 2012). However, there are significant differences between burning to benefit big game and their habitat and burning for other ecological factors, such as brush control, mimicking “natural” fire regimes, or urban-interface clearing. Optimal burning prescriptions for big game habitat differ from other burning prescriptions in terms of season of burn, intensity of burn, and other factors. Consequently, many burns, even if ostensibly conducted to benefit big game, actually have fewer positive benefits than they could have for big game. Hence, it is important to understand how to burn to maximize benefits for big game.
Prescribed burns for big game are most common in the understory (especially the herbaceous layer) of piñon (Pinus edulis)-juniper (Juniperus spp.) woodlands and other forest types, grasslands, and shrublands. The effects of fire vary with the intensity and timing of the burn (Whelan, 1995; Arno and Allison-Bunnell, 2002). For managing big game habitat, most prescribed fire is low-intensity because of the desired effects of burning on vegetation (see below), because cooler fires are promoted during the optimal burning periods, and because cooler burns minimize flame lengths and thus the danger of torching and transitioning into a crown fire (Whelan, 1995; Lutz et al., 2003; Fulé et al., 2004). Fire can significantly affect wildlife habitat, and the following sections generalize these effects on big game habitat.
Community structure. Fire has its greatest impact on the structure of plant communities, usually by delaying succession. It does this by (1) shifting communities from tree-dominated overstory to grass/forb or shrub communities, (2) decreasing dominance of one or a few climax herbaceous or woody species and thereby increasing forage diversity, (3) shifting shrubs from an older, decadent state to a younger state with less lignified growth and other anti-herbivory adaptations, and (4) eliminating dead aboveground biomass (Hobbs and Spowart, 1984; Whelan, 1995; Bender, in press). For example, fire can increase the quality of big game diets to a much greater extent than gains derived from increased nutrients in forages alone because it removes dead plant material and thus allows animals access to newly emergent forages in early phenological states (i.e., high in cell-soluble nutrients, low in cellulose and other structural carbohydrates) (Hobbs and Spowart, 1984). Removing cured aboveground biomass and thus exposing higher-quality forages also facilitates foraging by big game, decreasing the time they must spend searching for food. This is important because ruminants are limited in the time they can spend feeding (see below).
Plant biomass. Biomass increases after burning due to a combination of fertilization and partial to full opening of the tree or shrub overstory or removal of ground litter (Jones and DeByle, 1985; Whelan, 1995).
Green-up. Fire speeds green-up of vegetation in the spring, at least in the first year. This occurs because the blackened ash left after burning warms in the sun, creating a warmer microclimate for plants to germinate or initiate growth (Hobbs and Spowart, 1984).
Plant composition. Fire can change the composition of plant communities depending upon the intensity and frequency of burning (Whelan, 1995). Intense fires can eliminate entire overstory tree communities, while frequent low-intensity fires can change plant composition by eliminating less fire-tolerant species (shrubs, which store reserves above ground) in favor of more fire-tolerant species (perennial grasses, which store reserves below ground) (Jones and DeByle, 1985; Whelan, 1995). In grasslands, frequent fire can also cause species shifts away from perennial grasses to forbs (Ford and McPherson, 1996; Bender, in press). Forbs can also be preferentially increased in abundance by spring fires as compared to burning in other seasons (Brewer and Platt, 1994).
Fertilization and nutrients. Increases in understory biomass, improvement of diet quality beyond levels attributable to increased forage quality alone (Hobbs and Spowart, 1984), and effects on nutritional quality of plants are the three principal benefits of fire for big game. The primary effect of fire on nutrients is increased protein levels, and is caused by freeing the nitrogen (N) bound in litter, especially the high carbon:nitrogen ratio (C:N) litter of conifer-dominated communities (Bender, in press). In general, fire increases protein levels approximately 25%, although this can range as high as 145% (Bender, in press; L. Bender, unpublished data). Further, the effects of fire on protein can vary with several factors, including:
- Plant species. In general, grasses show the least response to fire, and shrubs the greatest. This is because grasses tend to store nutrients below ground, so burning the cured aboveground biomass frees few nutrients. Conversely, shrubs maintain much of their live tissue above ground, so burning potentially frees many more nutrients.
- N content of soils. If soils are high in N, gains from burning will be less. For vegetation types present in New Mexico, the greatest nutrient gains would occur in conifer-dominated systems (piñon-juniper and montane conifer, including ponderosa pine [Pinus ponderosa]) because of the high C:N ratio of their litter and consequent slow decomposition rates.
- Time since burn. Protein (and other nutrient) gains from burning are relatively short-term, generally lasting only 1 to 2 years. After 3 to 5 years, there is usually no detectable difference (Bender, in press). However, the total forage biomass response to burning lasts significantly longer, and only begins to change as the overstory canopy closes or ground cover of litter increases.
- Season of burn. Prescribed burns (or any other management actions) should provide needed nutrients when required by the target species. The season of burn is important because different seasons result in different times of increased protein levels in forage. In general, the relationships between season of burn and protein levels in plants are as follows (Einarsen, 1946; DeWitt and Derby, 1955; Grelen and Epps, 1967; Dills, 1970; Hobbs and Spowart, 1984; Bender, in press).
a. Winter burns. Result in protein increases in early spring; protein levels decline throughout the summer.
b. Spring burns. Result in protein increase in late spring and early summer; protein levels decline throughout mid- and late summer.
c. Summer burns. Result in a large increase in protein in mid-summer followed by a rapid decline in late summer.
d. Autumn burns. Generally result in little response because plants have little time to make a growth response. Autumn burns also pose the greatest threat of loss of nutrients due to runoff, leaching, and windblow because of the length of exposure before plants initiate growth in early spring.
Fire can also affect other nutrients. Fire tends to decrease crude fiber of plants, thus increasing digestibility. Because digestibility and digestible energy are synonymous, fire therefore results in increased energy available, both within plants (because of increased digestibility) and overall (because of increased plant biomass) (Edwards et al., 2004). Gains in digestibility tend to be longer-lived than protein gains, lasting as long as 10 years or until significant competition begins to cause declines in understory biomass. Fire has variable effects on mineral levels of forages, with spring burns tending to increase mineral levels and burns in other seasons having little effect. Fire has a small effect on fat levels (ether extracts) of plants; however, grasses in general tend to show increased ether extracts after burning.
For big game, the key periods of energy, mineral, and especially protein needs are late spring and summer, corresponding with the last trimester of gestation, lactation, and antler growth (Verme and Ullrey, 1984; Wakeling and Bender, 2003; Bender, 2018). During this period, demands for both protein and energy are greatest to invest in the growing fetus and to maintain lean muscle tissue or accumulate fat in the female. High protein content in milk is critical for rapid growth of juveniles (Landete-Castilljos et al., 2003) and maintenance of female body condition, both of which are critical for the survival of fawns and adults (Bender et al., 2007, 2011, 2012, 2013a; Lomas and Bender, 2007; Hoenes, 2008; Bender and Piasecke, 2010). Similarly, growing antlers are more than 80% protein, so bucks and bulls require a minimum of 16%, and ideally more than 20%, protein in their diets during the primary antler growth period from late April through early September (Bender, 2018). Increased mineral content also benefits antler development (Bender, 2018).
Late winter and spring burns (generally March to April, depending on the location in New Mexico) can provide a critical protein flush during this period, thereby increasing survival, productivity, and performance of big game. Additionally, spring burns can also increase mineral content of forages, providing minerals during the period of rapid antler development in males (Bender, 2018). Big game will also consume ash following a burn, which provides a further, direct source of minerals in their diets.
Later burns, as are typical of public lands in New Mexico (i.e., June to August) in the belief that lightning strikes determined presettlement or “natural” fire regimes (but see, for example, Williams, 2002), can still increase biomass of forage for big game, but much of the beneficial effects of burning on nutrient content and rapid green-up of forage are lost. Because the freeing of nutrients is probably the most important advantage of fire over mechanical, herbicidal, and livestock grazing treatments for big game, managers should burn early to maximize nutrient benefits if big game habitat is the primary reason for prescribed burns. Additionally, late winter and spring burns are cooler than later burns, resulting in less mortality of shrubs and remaining tree overstory, and patchier burns, which maintains critical forages (shrubs), cover (shrubs and trees), and ecotones (i.e., edges) while increasing quality and quantity of forages (Figure 1). Thus, burns aimed at increasing nutritional quality of big game ranges should be timed for late winter or early spring to increase availability of key nutrients for big game productivity at the critical time they are required.

Figure 1. Thorough herbaceous burn in short grassland. Burning in late winter will encourage production of forbs, which are important big game foods. Perennial grasses will remain, but their coverage will be decreased 10 to 15% to open space for forbs. This relatively cool burn maintained the juniper at the crest of the hill as cover.
One often overlooked benefit of burning is that burns can increase the foraging efficiency of big game because low-quality forage (such as cured grasses) is removed by the fire. Big game, especially deer and pronghorn, are very selective feeders because of their high dietary quality requirements. However, they are limited to about 12 to 13 hours per day that they can spend feeding because of the time required for rumination and the time required for foods to move through their complex digestive system (i.e., retention times), which increases as the quality of forage declines (Duncan et al., 1998; Holand et al., 1998; Cook, 2002). Thus, they have limited time to find the nutritious forage they need.
The higher-quality foods they require are also much rarer components of the foraging landscape (Hoenes and Bender, 2012). Moreover, because retention times increase as forage quality declines, big game cannot simply increase nutrient gains by eating more low-quality foods (Duncan et al., 1998; Holand et al., 1998; Cook, 2002; Figure 2). For example, handling times of low-quality foods (i.e., ca. 55% digestibility) are >30 hours for small deer, while handling times of high-quality foods (ca. 80% digestibility) are <20 hours (Holand et al., 1998). Deer thus retain more of the low-quality food in the stomach complex longer, which lessens feeding time because of the increased time needed for digestion (and decreases the amount of higher-quality food they can eat). So, the lower the forage quality or the longer the search times, the lower their nutrient gain. By removing low-quality forages such as cured grasses, fire increases the efficiency at which big game can locate preferred foods such as forbs. This results in decreased search times (which are longer as standing biomass increases; Figure 2) and thus better allows big game to meet both diet quality and quantity requirements in the time they have available to feed each day.
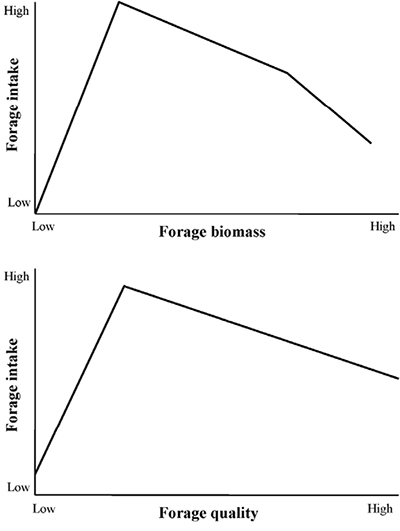
Figure 2. Simplified relationships between forage intake and forage quality and standing biomass. Wild herbivores cannot meet nutritional needs by increasing intake as forage quality declines because of longer handling times of lower-quality foods. Also, search times increase as standing biomass increases above ca. 1,000 pounds per acre, which also decreases intake because herbivores are limited in the time they have to forage. Optimal foraging occurs when forage quality is high and standing biomass is reduced, resulting in shorter search times and higher diet qualities. Burning can provide these conditions.
Fire behavior for big game burns
The exact fire behavior and burn plan are highly dependent on the local vegetation and topography, and thus need to be developed on a site-specific basis. In general, low humidity and a 5- to 10-mph wind provide ideal burning characteristics with 1- to 2-foot flame lengths under a range of temperatures (i.e., 50–90°F) and fine fuel moisture content down to ca. 2% (Figure 3) in arid and semi-arid New Mexico habitats. Where ground cover is extremely patchy, such as in desert grassland or in years of poor grass production or heavy grazing, winds may have to be stronger to get a fire to carry. Fire intensity, and thus flame lengths, may also need to be higher if a significant kill of tall shrubs (especially deciduous species) or overstory trees in piñon-juniper woodlands is a management objective (Figure 4). However, foliage moisture of conifers is usually low during late winter or early spring burns, so the difficulty is often keeping these conifers from burning to maintain adequate cover (Bender, in press). This is particularly true with juniper, which often shows a shrub-like growth form where the highly flammable foliage is in close or direct contact with the herbaceous ground cover. In these cases, woodlands may require fuel treatments to keep from having too much overstory kill when burning during these times (see below).
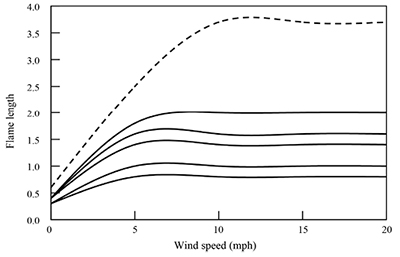
Figure 3. Predicted fire intensity in sparse to low-load, short herbaceous communities on White Sands Missile Range grasslands and woodland understories. In addition to wind speed, variables included slope, live herbaceous fuel moisture, and fine fuel moisture (simulated using fire models GR1, GR2, GS1, and TU1 in the program BehavePlus 5.0.5; Heinsch and Andrews, 2010). Fire intensity is fine for big game burns except for higher fuel loads under extremely dry, low-humidity conditions (i.e., fine fuel moisture <1%; black-dashed curve), where flame lengths could lead to significant torching of trees if present.

Figure 4. Even late winter burns in March can generate long flame lengths and intense fire. Managers need to tailor fire intensity to burn objectives—the greater the intensity, the higher the kill of woody species, which is counterproductive for many big game burns.
Fire intensity is also affected by the type of burn. Headfires (burning with the wind, i.e., in the direction the wind is blowing) will provide a fast, patchy burn that is ideal for minimizing mortality of woody species and creating a mosaic of burned and unburned patches, which is often optimal for big game habitat (Figure 5). Burning across the wind (flankfire) or into the wind (backfire) results in progressively slower, hotter, and less patchy burns. These burns will usually result in increased shrub and tree mortality. Similarly, upslope burns are usually faster and patchier than downslope burns.

Figure 5. Late winter burns are frequently patchy, resulting in a mosaic of burned and unburned areas. This increases the diversity of the rangeland and the variety of forage available to big game and livestock.
With regards to fire intensity, managers tend to overestimate flame lengths and thus intensity of burns. For example, they frequently mistake torching succulents or shrubs for the actual fire flame length. If unsure, fire intensity can also be determined after the burn by observing characteristics of remaining vegetation and the degree of charring on the ground. As noted above, ideal burns for big game are low-intensity burns, with flame lengths <3 feet and light ground char characterized by black ash in a mosaic of burned and unburned patches (Tables 1 and 2). Using post-burn assessments based on scorch or ground char criteria can allow managers lacking experience with prescribed fire to gain knowledge of fire behavior specific to the local characteristics of their ownerships, and thereby better tailor future burns to maximize benefits for big game.
|
Table 1. Generalized Criteria for Determining Fire Intensity (i.e., flame length) Based on Effects on Woody Vegetation (adapted from Ryan and Noste [1985]) |
||
|
Flame length (feet) |
Crown scorch height (feet) |
Sizes of trees killed (diameter at breast height in inches) |
|
0–2 |
0–9 |
Seedling (<1.0) |
|
2–4 |
9–24 |
Sapling (1.0–4.9) |
|
4–8 |
24–64 |
Pole (5.0–8.9) |
|
8–12 |
64–116 |
Small saw timber (9.0–13.0) |
|
>12 |
>116 |
Large saw timber (>13.0) |
|
Table 2. Simplified Criteria for Determining Surface Char Class in Grasslands, Shrublands, and Woodlands Following Fire (adapted from Ryan and Noste [1985] and Jain et al. [2008]; see Ryan and Noste [1985] for more inclusive characteristics) |
|
|
Surface char |
Site characteristics |
|
Uncharred |
No evidence of burning on the ground. Some vegetation may be scorched by heat. |
|
Light ground char |
Burn is irregular and spotty; >30–40% litter cover remains. Surface is predominantly black (black ash) immediately after the burn. Some gray ash may be present immediately after the fire, but soon becomes inconspicuous. Charring may extend slightly into the soil surface, but the soil is not otherwise altered. Some herbaceous plant parts (bases, stems, etc.) are still present. Some leaves and small twigs remain on shrubs. Less than 60% of the brush canopy is commonly consumed. |
|
Moderate ground char |
Litter is mostly consumed; <10% charred litter may remain. Surface is covered with gray or white ash immediately after the burn. Ash soon disappears, leaving bare mineral soil. Charring extends slightly into mineral soil, but the soil is not otherwise altered. Herbaceous plant parts totally consumed to ground level. Some charred stems remain on shrubs, generally >1/4- to 1/2-inch in diameter. 40–80% of the brush canopy is commonly consumed. May show lines of gray char from logs or other woody debris. |
|
Deep ground char |
Surface consists of fluffy white ash immediately after the burn. Ash soon disappears, All organic matter is consumed in the mineral soil to a depth of 1/2–1 inch. This is underlain Large branches are burned, and only stubs >1/2-inch in diameter remain. Orange-colored char or soils under logs, brush piles, etc., due to prolonged, high heat. |
Managers thus need to consider plant biomass (fuel load), topography, desired vegetation changes (for example, high or low shrub kill), and weather when planning their burns for big game habitat (Figure 6). Assistance in fire planning is available from private, county, state, and federal land management agencies.

Figure 6. A good big game burn in open piñon-juniper and oakbrush. The herbaceous layer is mostly burned but some patches remain, some of the oakbrush is topkilled and will re-sprout, and much of the scattered piñon-juniper will survive.
Burning Considerations in Selected Plant Communities
Some general considerations for burning to promote big game habitat in the more common plant communities of New Mexico are given in the following sections. A few issues are common to all communities, however. First, unless accompanied by forage seedings, understory responses will be limited to the species already present. If few high-quality forages are on the site, managers may want to consider seeding following burning but prior to the rainy season (Bender, 2012; Bender, in press). Additionally, noxious weeds are always a concern, especially where burning is severe or accompanied by significant soil disturbance. If noxious species are prevalent, seeding with high-quality forages is a good method of limiting their establishment.
Piñon-juniper woodland. Most forage benefits for big game in piñon-juniper come from increasing forage biomass and species diversity by opening the piñon-juniper overstory and/or by increasing forage quality of the understory in more open woodlands (Van Hooser et al., 1993; Bender, 2012; Bender, in press), but changes in big game use or big game forage following mechanical manipulation only (thinning, cabling, chaining, chipping, etc.) have been mixed or even counterproductive (Howard et al., 1987), especially where chipping or other mastication treatments leave the biomass in place (Bender, in press). This can be true even if such treatments are accompanied by seeding (Huffman et al., 2017). In contrast, big game consistently show positive responses to burning piñon-juniper (Greenwood et al., 1999; Erskine and Goodrich, 1999; Bender, in press). This is because conifer-dominated communities have litter with a high C:N ratio that results in slow rates of decomposition since microbial decomposers are limited by a lack of N to build microbial protein (i.e., bodies) (Miller et al., 1979). Consequently, most conifer systems are nutrient-deficient because many nutrients, especially N, are tied up in the trees or litter. By converting litter and tree biomass to ash, fire frees nutrients and makes them available to understory grasses, forbs, and shrubs (Miller et al., 1979; White, 1996; Whelan, 1995; Bender, in press), increasing forage quality. Along with opening the overstory, the combination of mineral soil, high nutrient availability, and sunlight provides ideal conditions for establishment of shade-intolerant species (Whelan, 1995; Arno and Allison-Bunnell, 2002; Schoennagel et al., 2004), which includes most preferred forages of big game.
Two important considerations in burning piñon-juniper are frequency of burning and intensity of burning. Longer fire intervals, on the range of 8 to 12 years, favor the establishment of shrubs in the understory. Conversely, shorter intervals of 3 to 6 years can eliminate shrubs and favor grasses and forbs (Bender, 2012). For big game species, such as deer and pronghorn, that require the highest-quality diets, shrubs are critical for maintaining adult survival in areas characterized by frequent drought, such as most of New Mexico (Bender, 2012; Bender et al., 2011, 2013b; Hoenes and Bender, 2012). In these cases, longer fire intervals should be favored to maintain and enhance shrub communities. Similarly, smaller big game species, such as mule deer, depend on significant hiding cover, and all big game will use overstory thermal cover in New Mexico during the hotter months (Bender et al., 2011, 2012, 2013b; Bender, in press). Consequently, it is not desirable to eliminate all overstory piñon-juniper (or even a substantial portion of it in certain areas), so cooler late-winter to early spring fires that limit fire intensity should be used to maintain cover as well as to maximize nutrient benefits of burning. However, because foliage moisture may be very low during these times (especially in late winter), fuel treatments may be necessary to limit scorching and tree mortality. These may include mowing around denser patches (or even individual trees with ground-level branches) that a manager wants to preserve for cover, eliminating high fuel loads under trees (such as pack rat middens), or burning when some residual snow cover remains under trees due to shading. Higher winds also move ground fires faster, limiting ignition of trees.
Montane conifer and ponderosa pine. Overstory canopy coverage is similarly the single most important variable in determining the biomass and composition of understory communities in montane conifer types (Pieper, 1990; Canham et al., 1994; Thomas et al., 1999). Fire can work in combination with thinning to greatly enhance the quantity and quality of understory forage in montane conifer types (Oswald and Covington, 1983; Covington et al., 1997). The effects of fire on forage are identical to those described under piñon-juniper, except that understory responses tend to be greater because montane conifer tends to have greater canopy coverage and occurs in more mesic (higher moisture level) sites, providing more moisture for understory responses. Thinning and prescribed burning have consistently resulted in significant increases in elk and deer use of montane stands (Lowe et al., 1978; Roberts and Tiller, 1985; Crouch, 1986). Burn intervals need to be adjusted as described under piñon-juniper to favor shrubs or grasses, depending on management goals, but burning intervals in montane conifer should be at the shorter end of the ranges listed, whereas most piñon-juniper sites should be burned at the longer intervals in the ranges listed. Residual overstory trees also suffer less mortality than residual juniper during follow-up burns because most montane conifer species have tree-like rather than shrub-like growth forms, and many are fire-adapted (e.g., ponderosa pine).
Aspen (Populus tremuloides). In higher-elevation mesic sites in New Mexico, remnant aspen stands provide extremely high-quality big game habitat. Aspen is shade-intolerant, so regeneration requires complete removal of the overstory (Bartos, 2001). In the past, this naturally occurred through stand-replacing wildfire, but more recently is a result of patch- or clear-cutting. If stands are cut, aspen regenerates much more vigorously if burned after cutting (Shepperd, 2001) due to near complete elimination of auxins (a class of plant hormone), which inhibit suckering (vegetative regrowth). Also, if a shade-tolerant conifer understory is present in mature aspen stands, conifers need to be killed by cutting or burning to regenerate aspen (Patton and Jones, 1977). Aspen reproduces poorly (if at all) from seed in the Southwest because of a lack of suitable soil characteristics and adequate moisture (Shepperd, 2001).
Because aspen is short-lived (100 to 125 years), many of the stands that are successionally changing to Douglas-fir (Pseudotsuga menziesii) and spruce (Picea spp.)/fir (Abies spp.) communities due to fire exclusion after approximately 1900 are in danger of being lost permanently. Aspen has declined in New Mexico from approximately 1.1 million acres historically to approximately 140,000 acres currently (Bartos, 2001). While burning can most effectively regenerate aspen, the challenge is treating enough acreage to allow adequate regeneration without losing small treatments to ungulate browsing (Shepperd, 2001; Halbritter and Bender 2011, 2015). For production of browse for deer and elk, an optimal rotation age is 20 to 30 years for aspen (Patton and Jones, 1977).
Short grasslands. Fire (along with grazing) is a predominant force in grassland management (Ford and McPherson, 1996; Allison and Bender, 2017). In general, fire results in a short-term (1 to 3 years) decrease in overall production of short grasslands, although this effect varies with precipitation; if precipitation is normal or above normal, the effect may be absent or last approximately 1 year, but if precipitation is below normal the effect may last longer (Launchbaugh, 1964; Wright and Bailey, 1980; Ford and McPherson, 1996). Burning increases forb density and diversity in short grasslands (Bailey and Anderson, 1978; Collins and Barber, 1986; Ford and McPherson, 1996; Bender, in press), enhancing forage quality for big game, which make limited use of grasses (excluding elk) but forage heavily on forbs when available (Pederson and Harper, 1978; Wakeling and Bender, 2003; Hoenes and Bender, 2012).
For short grasslands, spring burns on a 3- to 6-year interval can facilitate an ideal balance of forbs and grasses, and along with fertilization effects can significantly enhance big game habitat quality. However, the short-term decrease in perennial grasses may lower grazing capacity for cattle. In desert grasslands, burn intervals are generally much longer and depend on the rate of plant recovery and production following burning. In more productive or ungrazed grasslands, live grass and other herbaceous cover may decline in the years following burning as litter increases (Figure 7), so ungrazed grasslands may require more frequent fire intervals.
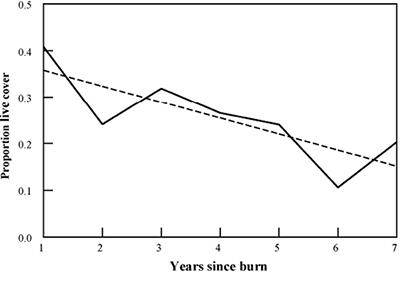
Figure 7. In ungrazed or highly productive grasslands, cover of live herbaceous forage can decrease in the years following burning because of accumulation of herbaceous plant litter. Burning on a <6-year interval can keep cover of live herbaceous forage high.
Fire can reduce woody plant cover (Ford and McPherson, 1996), and because short grasslands often lack cover for species such as mule deer, shortgrass prairie sites with woody cover (or where woody cover is being established) should be burned with very low-intensity spring fires at longer intervals of 8 to 10 years or protected until shrubs are established.
Shrublands. Oakbrush (Quercus spp.) and true mountain mahogany (Cercocarpus montanus) are the shrubland vegetation most benefitted by burning in New Mexico, although bitterbrush (Purshia tridentata) and sagebrush (Artemisia spp.; most commonly A. tridentata and A. filifolia) can also benefit from burning. Because shrubs store reserves above ground, fire frequencies need to be longer to avoid stressing or eliminating shrubs, and burns should be in dormant seasons to minimize mortality as well as provide the optimal nutritional benefits for big game. In general, burn intervals of 8 to 10 years are ideal for rejuvenating shrublands as well as minimizing the density of regeneration of sprouting species after fire. With many browse species, re-sprouting after burning can be extremely dense; however, seeding with herbaceous species after burning or using mechanical disturbance (or livestock grazing) between fires can decrease the density of sprouts, resulting in more open stands with a more diverse and productive herbaceous understory (Stevens and Davis, 1985). Such a treatment can greatly enhance the overall quality of these areas for big game.
Oakbrush is the most widely distributed shrub community in New Mexico, and oakbrush is an extremely aggressive re-sprouter from root crowns following burning or other disturbance (Harrington, 1985). Oakbrush should be burned in winter or early spring, which does not harm the clones and encourages re-sprouting; hot summer burns can inhibit regrowth and may kill clones (Harrington, 1985). When rejuvenated by burning, big game use of oakbrush greatly increases (Stevens and Davis, 1985); in contrast to mountain mahogany, oakbrush browse is only moderately palatable, and young shoots and new growth are the most preferred portions of the shrubs, likely because rapidly growing new growth contains fewer tannins (James et al., 1980).
For species such as sagebrush and bitterbrush that are more vulnerable to fire-induced mortality, burning should be limited to dormant season fires, particularly in late winter (Stevens and Monsen, 2004), and fire intervals may need to be extended to 20 years or more, depending on the level of fire-induced mortality. Sagebrush in particular is easily killed by fire, so burned areas should be relatively small, resulting in a mix of sage and herbaceous plants. Fire intervals should be timed to the closing of the sagebrush canopy; ideal cover is less than 50% sagebrush. In areas such as northwestern New Mexico where cheatgrass (Bromus tectorum) is common, additional actions, such as seeding with desirable grasses and forbs, may be necessary to prevent the burned areas from becoming dominated by this invasive, exotic
annual grass.
Literature Cited
Allison, C.D., and L.C. Bender. 2017. Grazing and biodiversity [Circular 686]. Las Cruces: New Mexico State University Cooperative Extension Service. https://pubs.nmsu.edu/_circulars/CR686/
Arno, S.F., and S. Allison-Bunnell. 2002. Flames in our forest: Disaster or renewal? Washington, D.C.: Island Press.
Bailey, A.W., and M.L. Anderson. 1978. Prescribed burning of a Festuca-Stipa grassland. Journal of Range Management, 31, 446–449.
Bartos, D.L. 2001. Landscape dynamics of aspen and conifer forests. In W.D. Shepperd, D. Binkley, D.L. Bartos, T.J. Stohlgren, and L.G. Eskew (Compilers), Sustaining aspen in western landscapes: Symposium proceedings (pp. 5–14) [United States Forest Service Proceedings RMRS-P-18]. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station.
Bender, L.C. In press. Elk, deer, and pinyon-juniper: Needs, what works, and what doesn’t. In Piñon-juniper habitats: Status and management for wildlife. Proceedings of a symposium, 2016 October 12–14, Albuquerque, NM. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station.
Bender, L.C. 2012. Guidelines for management of habitat for mule deer: Piñon-juniper, Chihuahuan desert, arid grasslands, and associated arid habitat types [Circular 662]. Las Cruces: New Mexico State University Cooperative Extension Service. https://pubs.nmsu.edu/_circulars/CR662/
Bender, L.C. 2018. Basics of trophy management [Guide L-111]. Las Cruces: New Mexico State University Cooperative Extension Service.
Bender, L.C., and J.R. Piasecke. 2010. Population demographics and dynamics of colonizing elk in a desert grassland-scrubland. Journal of Fish and Wildlife Management, 1, 152–160.
Bender, L.C., R.A. Baldwin, and J.R. Piasecke. 2012. Habitats occupied by elk (Cervus elaphus) in desert grassland–scrublands of northwestern New Mexico. The Southwestern Naturalist, 57, 361–369.
Bender, L.C., B.D. Hoenes, and C.L. Rodden. 2012. Factors influencing survival of desert mule deer in the greater San Andres Mountains, New Mexico. Human–Wildlife Interactions, 6, 245–260.
Bender, L.C., L.A. Lomas, and J. Browning. 2007. Condition, survival, and cause-specific mortality of adult female mule deer in north-central New Mexico. Journal of Wildlife Management, 71, 1,118–1,124.
Bender, L.C., J.C. Boren, H. Halbritter, and S. Cox. 2011. Condition, survival, and productivity of mule deer in semiarid grassland-woodland in east-central New Mexico. Human–Wildlife Interactions, 5, 276–286.
Bender, L.C., J.C. Boren, H. Halbritter, and S. Cox. 2013a. Factors influencing survival and productivity of pronghorn in a semiarid grassland-woodland in east-central New Mexico. Human–Wildlife Interactions, 7, 313–324.
Bender, L.C., J.C. Boren, H. Halbritter, and S. Cox. 2013b. Effects of site characteristics, pinyon-juniper control, and precipitation on habitat quality for mule deer in New Mexico. Human–Wildlife Interactions, 7, 47–59.
Brewer, J.S., and W.J. Platt. 1994. Effects of fire season and herbivory on reproductive success of a clonal forb, Pityopsis graminifolia. Journal of Ecology, 82, 665–675.
Canham, C.D., A.C. Finzi, S.W. Pacala, and D.H. Burbank. 1994. Causes and consequences of resource heterogeneity in forests: Interspecific variation in light transmission by canopy trees. Canadian Journal of Forest Research, 24, 337–349.
Collins, S.L., and S.C. Barber. 1986. Effects of disturbance on diversity in mixed-grass prairie. Vegetatio, 64, 87–94.
Covington, W.W., P.Z. Fulé, M.M. Moore, S.C. Hart, T.E. Kolb, J.N. Mast, S.S. Sackett, and M.R. Wagner. 1997. Restoring ecosystem health in ponderosa pine forests of the Southwest. Journal of Forestry, 95(4), 23–29.
Crouch, G.L. 1986. Effects of thinning pole-sized lodgepole pine on understory vegetation and large herbivore activity in central Colorado [Research Paper RM-268]. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station.
DeWitt, J.B., and J.V. Derby, Jr. 1955. Changes in nutritive value of browse plants following forest fires. Journal of Wildlife Management, 19, 65–70.
Dills, G.G. 1970. Effects of prescribed burning on deer browse. Journal of Wildlife Management, 34, 540–545.
Edwards, S.L., S. Demarais, B. Watkins, and B.K. Strickland. 2004. White-tailed deer forage production in managed and unmanaged pine stands and summer food plots in Mississippi. Wildlife Society Bulletin, 32, 739–745.
Einarsen, A.S. 1946. Crude protein determination of deer food as an applied management technique. Transactions of the North American Wildlife Conference, 11, 309–312.
Erskine, I., and S. Goodrich. 1999. Applying fire to pinyon-juniper communities of the Green River corridor, Daggett County, Utah. In S.B. Monsen and R. Stevens (Compilers), Proceedings: Ecology and management of pinyon-juniper communities within the interior west ; 1997 September 15-18; Provo, UT (pp. 315–316) [Proceedings RMRS-P-9]. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station.
Ford, P.L., and G.R. McPherson. 1996. Ecology of fire in shortgrass prairie in the southern Great Plains. In D.M Finch (Ed.), Ecosystem disturbance and wildlife conservation in western grasslands - A symposium proceedings, September 22-26, 1994; Albuquerque, NM (pp. 20–39) [General Technical Report RM-GTR-285]. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station.
Fulé, P.Z., A.E. Cocke, T.A. Heinlein, and W.W. Covington. 2004. Effects of an intense prescribed forest fire: Is it ecological restoration? Restoration Ecology, 12, 220–230.
Greenwood, C.L., S. Goodrich, and J.A. Lytle. 1999. Response of bighorn sheep to pinyon-juniper burning along the Green River corridor, Daggett County, Utah. In S.B. Monsen and R. Stevens (Compilers), Proceedings: Ecology and management of pinyon-juniper communities within the interior west ; 1997 September 15-18; Provo, UT (pp. 205–209) [Proceedings RMRS-P-9]. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station.
Grelen, H.E., and E.A. Epps, Jr. 1967. Season of burning affects herbage quality and yield on pine-bluestem range. Journal of Range Management, 20, 31–33.
Halbritter, H., and L.C. Bender. 2011. Quality of habitat occupied by elk (Cervus elaphus) in the southern Sacramento Mountains, New Mexico. The Southwestern Naturalist, 56, 1–8.
Halbritter, H.H., and L.C. Bender. 2015. Herbivory of sympatric elk and cattle on Lincoln National Forest, south-central New Mexico. Forest Ecosystems, 2, 25.
Harrington, M.G. 1985. Notes: The effects of spring, summer, and fall burning on Gambel oak in a southwestern ponderosa pine stand. Forest Science, 31, 156–163.
Heinsch, F.A., and P.L. Andrews. 2010. BehavePlus fire modeling system, version 5.0: Design and features [General Technical Report RMRS-GTR-249]. Ft. Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station.
Hobbs, N.T., and R.A. Spowart. 1984. Effects of prescribed fire on nutrition of mountain sheep and mule deer during winter and spring. Journal of Wildlife Management, 48, 551–560.
Hoenes, B. 2008. Identification of factors limiting desert mule deer populations in the greater San Andres Mountains of southcentral New Mexico [Master’s thesis]. Las Cruces: New Mexico State University.
Hoenes, B.D., and L.C. Bender. 2012. Factors influencing foraging habitats of mule deer (Odocoileus hemionus) in the San Andres Mountains, New Mexico. The Southwestern Naturalist, 57, 370–379.
Howard, Jr., V.W., K.M. Cheap, R.H. Hier, T.G. Thompson, and J.A. Dimas. 1987. Effects of cabling pinyon-juniper on mule deer and lagomorph use. Wildlife Society Bulletin, 15, 242–247.
Huffman, D.W., M.T. Stoddard, J.D. Springer, and J.E. Crouse. 2017. Understory responses to tree thinning and seeding indicate stability of degraded pinyon-juniper woodlands. Rangeland Ecology and Management, 70, 484–492.
Jain, T.B., W.A. Gould, R.T. Graham, D.S. Pilliod, L.B. Lentile, and G. Gonzalez. 2008. A soil burn severity index for understanding soil-fire relations in tropical forests. AMBIO, 37, 563–568.
James, L.F., R.F. Keeler, A.E. Johnson, M.C. Williams, E.H. Cronin, and J.D. Olsen. 1980. Plants poisonous to livestock in the western states [Agriculture Information Bulletin 415]. Washington, D.C.: U.S. Department of Agriculture.
Jones, J.R., and N.V. DeByle. 1985. Fire. In N.V. DeByle and R.P. Winokur (Eds.), Aspen: Ecology and management in the western United States (pp. 77–81) [General Technical Report RM-119]. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station.
Landete-Castillejos, T., A. García, J.A. Gómez, and L. Gallego. 2003. Lactation under food constraints in Iberian red deer Cervus elaphus hispanicus. Wildlife Biology, 9, 131–139.
Launchbaugh, J.L. 1964. Effects of early spring burning on yields of native vegetation. Journal of Range Management, 17, 5–6.
Lomas, L.A., and L.C. Bender. 2007. Survival and cause-specific mortality of neonatal mule deer fawns, north-central New Mexico. Journal of Wildlife Management, 71, 884–894.
Lowe, P.O., P.F. Ffolliott, J.H. Dietrich, and D.R. Patton. 1978. Determining potential wildlife benefits from wildfire in Arizona ponderosa pine forests [General Technical Report RM-52]. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station.
Lutz, D.W., B.F. Wakeling, L.H. Carpenter, D. Stroud, M. Cox, D. McWhirter, S. Rosenstock, L.C. Bender, and A.F. Reeve. 2003. Impacts and changes to mule deer habitat. In J.C. deVos, Jr., M.R. Conover, and N.E. Headrick (Eds.), Mule deer conservation: Issues and management strategies (pp. 13–61). Logan: Berryman Institute Press, Utah State University.
Miller, H.G., J.M. Cooper, J.D. Miller, and O.J.L. Pauline. 1979. Nutrient cycles in pine and their adaptation to poor soils. Canadian Journal of Forest Research, 9, 19–26.
Oswald, B.P., and W.W. Covington. 1983. Changes in understory production following a wildfire in southwestern ponderosa pine. Journal of Range Management, 36, 507–509.
Patton, D.R., and J.R. Jones. 1977. Managing aspen for wildlife in the Southwest [General Technical Report RM-37]. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station.
Pederson, J.C., and K.T. Harper. 1978. Factors influencing productivity of two mule deer herds in Utah. Journal of Range Management, 31, 105–110.
Pieper, R.D. 1990. Overstory-understory relations in pinyon-juniper woodlands in New Mexico. Journal of Range Management, 43, 413–415.
Roberts, T.A., and R.L. Tiller. 1985. Mule deer and cattle responses to a prescribed burn. Wildlife Society Bulletin, 13, 248–252.
Ryan, K.C., and N.V. Noste. 1985. Evaluating prescribed fires. In J.E. Lotan, B.M. Kilgore, W.C. Fischer, and R.W. Mutch (Tech. Coords.), Proceedings—Symposium and workshop on wilderness fire (pp. 230–238) [General Technical Report INT-182]. Ogden UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station.
Schoennagel, T., T.T. Veblen, and W.H. Romme. 2004. The interaction of fire, fuels, and climate across Rocky Mountain forests. BioScience, 54, 661–676.
Shepperd, W.D. 2001. Manipulations to regenerate aspen ecosystems. In W.D. Shepperd, D. Binkley, D.L. Bartos, T.J. Stohlgren, and L.G. Eskew (Compilers), Sustaining aspen in western landscapes: Symposium proceedings (pp. 355–365) [Proceedings RMRS-P-18]. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station.
Stevens, R., and J.N. Davis. 1985. Opportunities for improving forage production in the Gambel oak types of Utah. In K.L. Johnson (Ed.), Proceedings of the third Utah shrub ecology workshop, 30–31 August 1983 (pp. 37–41). Logan: College of Natural Resources, Utah State University.
Stevens, R., and S.B. Monsen. 2004. Guidelines for restoration and rehabilitation of principal plant communities. In S.B. Monsen, R. Stevens, and N.L. Shaw (Compilers), Restoring western ranges and wildlands, vol. 1 (pp. 199–294) [General Technical Report RMRS-GTR-136-vol-1]. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station.
Thomas, S.C., C.B. Halpern, D.A. Falk, D.A. Liguori, and K.A. Austin. 1999. Plant diversity in managed forests: Understory responses to thinning and fertilization. Ecological Applications, 9, 864–879.
Van Hooser, D.D., R.A. O’Brien, and D.C. Collins. 1993. New Mexico’s forest resources [Resource Bulletin INT-79]. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station.
Verme, L.J., and D.E. Ullrey. 1984. Physiology and nutrition. In L.K. Halls (Ed.), White-tailed deer: Ecology and management (pp. 91–118). Harrisburg, PA: Stackpole Books.
Wakeling, B.F., and L.C. Bender. 2003. Influence of nutrition on mule deer biology and ecology. In J.C. deVos, Jr., M.R. Conover, and N.E. Headrick (Eds.), Mule deer conservation: Issues and management strategies (pp. 93–118). Logan: Berryman Institute Press, Utah State University.
Whelan, R.J. 1995. The ecology of fire. Cambridge, UK: Cambridge University Press.
White, C.S. 1996. The effects of fire on nitrogen cycling processes within Bandelier National Monument, NM. In C.D. Allen (Ed.), Fire effects in southwestern forests: Proceedings of the second La Mesa fire symposium (pp. 123–139) [General Technical Report RM-GTR-286]. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station.
Williams, G.W. 2002. Aboriginal use of fire: Are there any “natural” plant communities? In C.E. Kay and R.T. Simmons (Eds.), Wilderness & political ecology: Aboriginal influences & the original state of nature (pp. 179–214). Salt Lake City: University of Utah Press.
Wright, H.A., and A.W. Bailey. 1980. Fire ecology and prescribed burning in the Great Plains—A research review [General Technical Report INT-77]. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station.
Acknowledgments. I thank C. Allison, Linebery Policy Center for Natural Resource Management; R. Baldwin, University of California, Davis; J. Boren, New Mexico State University; A. Darrow, Mexico Coast Land and Cattle; B. Hoenes, Washington Department of Fish and Wildlife; and O. Rosas-Rosas, Colegio de Postgraduados, San Luis Potosí, for reviewing this publication.
For further reading
CR-662: Guidelines for Management of Habitat for Mule Deer
https://pubs.nmsu.edu/_circulars/CR662/
CR-664: Guidelines for Monitoring Elk and Mule Deer Numbers in New Mexico
https://pubs.nmsu.edu/_circulars/CR664/
CR-669: Using Population Ratios for Managing Big Game
https://pubs.nmsu.edu/_circulars/CR669/

Lou Bender is a Research Scientist (Wildlife) with the Department of Extension Animal Sciences and Natural Resources at NMSU. He earned his Ph.D. from Michigan State University. His research and management programs emphasize ungulate and carnivore management, integrated wildlife and livestock habitat management, and wildlife enterprises in the Southwest and internationally.
To find more resources for your business, home, or family, visit the College of Agricultural, Consumer and Environmental Sciences on the World Wide Web at pubs.nmsu.edu
Contents of publications may be freely reproduced for educational purposes. All other rights reserved. For permission to use publications for other purposes, contact pubs@nmsu.edu or the authors listed on the publication.
New Mexico State University is an equal opportunity/affirmative action employer and educator. NMSU and the U.S. Department of Agriculture cooperating.
Revised April 2018 Las Cruces, NM