Reproductive Tract Anatomy and Physiology of the Cow
Guide B-212
Reviewed by Jason Turner
College of Agricultural, Consumer and Environmental Sciences
Author: Professor/Extension Horse Specialist, Department of Extension Animal Sciences and Natural Resources, New Mexico State University (Print-friendly PDF)
Introduction
Understanding the anatomy and physiology of the cow's reproductive system is fundamental to good cattle management. Basic knowledge in this area will help producers do a better job of getting cows rebred, especially when using artificial insemination and estrus synchronization. It will also enable producers to better understand and control reproductive diseases and calving problems.
Anatomy
The ovary is the primary female reproductive organ and has two important functions: producing the female reproductive cell (the egg or ovum) and producing the hormones estrogen and progesterone. The cow's two ovaries are oval to bean-shaped organs that are 1—1.5 inches long and located in the abdominal cavity.
The secondary sex organs are a series of tubes that receive semen, transport sperm to the egg so it can be fertilized, nourish the fertilized egg (embryo), and allow the calf to be birthed. These organs include the vagina, cervix, uterus, uterine horns, and oviducts (also called Fallopian tubes), which each have a funnel-shaped opening called the infundibulum. Figure 1 presents a diagram of the complete reproductive tract anatomy.
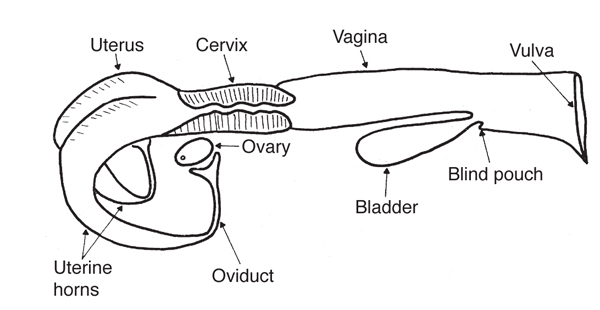
Figure 1. Diagram of the reproductive tract of the cow. (Adapted from Rich and Turman, n.d.)
The ovary produces the egg by a process called oogenesis. In contrast to the continuous production of sperm (spermatogenesis) in the male, oogenesis is cyclic. This cycle (called the estrous cycle) has a characteristic length and consists of a definite sequence of events, both physiological and behavioral.
The ovary contains several thousand tiny structures called primary follicles. Each follicle consists of a germ cell surrounded by a layer of cells. This germ cell has the potential to mature into an egg if the follicle completes development. However, most of the primary follicles never develop. Rather, they die, are absorbed by the ovary, and are replaced by newly formed primary follicles.
The relatively few primary follicles that develop completely do so through a series of phases. Many layers of cells are added to the single layer of cells surrounding the egg in the primary follicle, forming a central cavity. As the follicle and cavity grow larger, the egg becomes attached to the back of the follicle (opposite the ovulation site) by a stalk of cells. As the follicle continues to grow rapidly, the side opposite the egg bulges from the surface of the ovary and becomes very thin. Once the follicle reaches this mature state it is called a Graafian follicle. At ovulation, the thin portion ruptures to release the contents of the follicle, including the egg.
Following ovulation, the cells that developed within the follicle differentiate to form the corpus luteum, which has the important function of producing progesterone.
The released egg is caught by the infundibulum and moves into the oviduct, where fertilization occurs if viable sperm are present. The egg remains capable of fertilization for only a few hours. Thus, it is very important that fertile sperm be present near the time of ovulation. The egg moves through the oviduct and into the uterus within the next three to four days. If it is fertilized, it begins embryological development in the uterus. If it is not fertilized, it degenerates.
The body of the uterus is relatively short and poorly developed, while the uterine horns are longer and well developed. The embryo develops in the uterine horns. The fetus develops within a layer of membranes called the placenta. The mother nourishes the fetus via diffusion of nutrients across the placenta. There is no direct blood connection between fetus and mother.
The cervix is essentially the neck of the uterus. It has thick walls and a small opening that is difficult to penetrate because of overlapping or interlocking folds (annular rings). It serves as a passageway for sperm deposited in the vagina and for the fetus at the time of birth. During pregnancy, it is usually filled with a thick secretion that serves as a plug to protect the uterus from infective material entering through the vagina.
The vagina serves as the receptacle for the male penis during copulation. During natural mating, semen is deposited in the vagina near the cervix. The urinary bladder opens to the exterior through the urethra, which opens into the vagina. This region of the vagina is restricted in size because of sphincter muscles associated with the urethral opening. The region behind the external urethral orifice is called the vestibule and is a common passageway for both the urinary and the reproductive systems. The external opening of the vagina is called the vulva.
Hormonal Regulation of the Female Reproductive Tract
Normal reproduction in the female depends on hormones, which are specific chemical substances produced by specialized glands called endocrine glands. These secretions pass into the blood and lymph fluids and are transported to various parts of the body, where they exert several specific effects. Reproductive hormones may originate in the hypothalamus, pituitary, gonads, uterus, or placenta.
Hypothalamic hormones are produced in the hypothalamus, a portion of the brain that is the neural control center for reproductive hormones. The role of hypothalamic hormones is to regulate the release of other hormones from the pituitary gland, and many hypothalamic hormones are therefore called releasing hormones. The primary releasing hormone of reproduction is gonadotropin-releasing hormone (GnRH). GnRH controls the release of follicle-stimulating hormone (FSH) and luteinizing hormone (LH) from the pituitary gland, located at the base of the brain. These pituitary hormones regulate the production of estrogen and progesterone from the ovary. FSH stimulates the follicle's growth, development, and function, while LH causes the follicle to rupture and the corpus luteum to develop.
The female hormone estrogen is produced by the Graafian follicle on the ovary. Another hormone originating from the ovary is progesterone, which is produced by the corpus luteum. Each has an important role in the female reproductive process.
Estrogen regulates several functions: the development and functioning of the secondary sex organs; the onset of heat, or estrus (the period of sexual receptivity); the rate and type of body tissue growth, especially fat deposition; and priming or preparing of the prepubertal heifer and postpartum cow for the onset of sexual activity.
Progesterone is the hormone of pregnancy. It suppresses the further development of follicles and estrogen secretion. While progesterone is being produced, the female does not exhibit estrus. Progesterone is necessary for preparing the uterus to receive the fertilized egg and maintains the proper uterine environment for pregnancy. Estrogen and progesterone are not completely separate in their effects because both are necessary for complete development of some important organs. Uterine development is initiated by estrogen and completed by progesterone. The fertilized egg will not attach and survive in the uterus unless that tissue has been properly prepared by the action of estrogen and progesterone. Estrogen causes rhythmic contractions of the uterus. On the other hand, progesterone has a quieting effect on the uterus, so there are no contractions that might disturb pregnancy.
Complete development of the mammary gland also depends on both hormones. Estrogen promotes the growth of the duct system, and progesterone is necessary for the development of the clusters of milk-secreting alveoli on the ducts. In general, estrogen makes things happen and progesterone calms them down.
A number of other hormones and hormone-like chemicals are known to have important roles in regulating the cow's reproductive system. Prostaglandins are secreted by many body tissues and perform a variety of functions. The prostaglandin primarily affecting the cow's estrous cycle is prostaglandin F2α (PGF2α), which is produced by the uterus. PGF2α is the natural luteolytic agent that causes the corpus luteum to regress late in the estrous cycle and allows a new estrous cycle to be initiated in the nonpregnant female. In a pregnant cow, a signal is sent from the developing embryo to the uterus to prevent PGF2α release, which allows the corpus luteum to persist throughout pregnancy. Maternal recognition of pregnancy is believed to occur between days 16 and 17 after fertilization. Injecting cows or heifers with PGF2α between days 6 and 16 of the estrous cycle will cause premature regression of the corpus luteum, with the best results achieved among females injected on days 10 to 16.
Injecting PGF2α during the first five (1—5) and last five (17—21) days of the estrous cycle will generally not cause luteal regression. The luteolytic response allows the use of PGF2α in estrus synchronization programs in cow herds and initiates abortion in feedlot heifers. Estrogens, progestins, prostaglandins, and GnRH have all been used in various combinations in effective estrus synchronization programs.
The Estrous Cycle
The cow's reproductive cycle consists of a series of events that occur in a definite order over a period of days. The estrous cycle in the cow averages 21 days (range is 17—24). During this time, the reproductive tract is prepared for estrus or heat (the period of sexual receptivity) and ovulation (egg release). Figures 2 and 3 outline the sequence of anatomical and hormonal changes that occur during a typical 21-day cycle in which pregnancy does not occur.
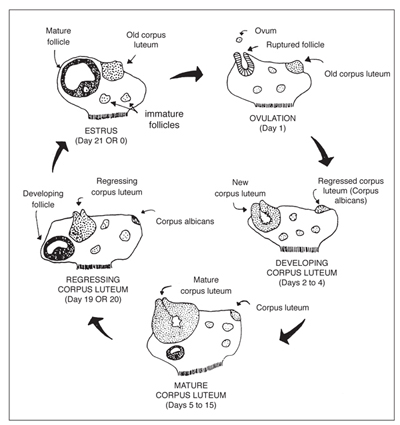
Figure 2. Diagram of a 21-day estrous cycle in which pregnancy does not occur. (Adapted from Deutscher, 1980)
Day 0: The cow is in estrus (standing heat) due to an increased concentration of estrogen. As estrogen levels reach a certain threshold level, a surge of LH is released by the pituitary. Near the end of standing heat, the mature Graafian follicle ovulates (ruptures) in response to this LH surge.
Days 1—2: The cells that formerly lined the follicle change and become the luteal cells of the corpus luteum. This change in cell form is caused by hormonal action, primarily the action of LH.
Days 2—5: The corpus luteum grows rapidly in both size and function. At this stage, numerous follicles may be seen on the ovary, but by day 5 they have begun to regress.
Days 5—16: The corpus luteum continues to develop and typically reaches its maximum growth and function by day 15 or 16. It secretes the hormone progesterone, which inhibits (blocks) LH release by the pituitary gland. During this period, the ovaries are relatively inactive except for the functional corpus luteum. No follicles reach maturity and/or ovulate because of high concentrations of progesterone.
Days 16—18: Increased follicular growth and accompanying estrogen secretion by the ovary stimulate PGF2αsecretion by the uterus, causing rapid regression of the corpus luteum.
Days 18—19: The corpus luteum is almost nonfunctional and progesterone release is suppressed, removing the blocking action of progesterone on LH and FSH. Of the several follicles that are initially recruited, one becomes dominant by a surge in rapid growth and activity. As this Graafian follicle grows, it secretes increasing amounts of estrogen, and the smaller follicles regress.
Days 19—20: With the increase in estrogen release by the Graafian follicle and a corresponding decrease in progesterone by the regressing corpus luteum, estrus or heat will occur (cycle has now returned to day 0). The high estrogen concentration in the blood triggers a release of LH near the onset of heat. Following this surge in LH blood concentrations, the mature follicle ruptures to release the egg, and the cellular tissue left behind becomes luteinized and forms a new corpus luteum (cycle has now returned to days 1—2). Progesterone again becomes the dominant hormone.
The timing given for these events is only approximate (based on a 21-day average) and differs for different cycle lengths (range of 17—24 days).
This discussion of events that occur during the estrous cycle is based on a full cycle in which pregnancy does not occur. If the egg is fertilized and begins developing in the uterus, the corpus luteum does not regress but continues to function and secrete progesterone. During pregnancy, no follicles develop to maturity and heat does not normally occur. Increased concentrations of progesterone promote uterine quiescence, providing the most favorable conditions for the developing fetus.
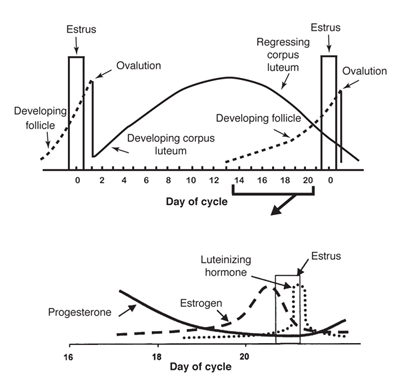
Figure 3. Graph of the anatomical and hormonal changes that occur during a typical 21-day estrous cycle. (Adapted from Deutscher, 1980)
Any condition that prolongs the period of time when blood concentrations of progesterone remain high (such as implanting, injecting, or feeding progestins for estrus synchronization) will keep the female from exhibiting estrus. Occasionally, the corpus luteum does not regress normally even though the animal does not become pregnant (pseudopregnancy).
Occasionally, abnormally short estrous cycles (7—11 days) occur. This condition appears to occur because either no corpus luteum is formed or, if one is formed, it is nonfunctional and progesterone concentration remains too low to sufficiently suppress the release of gonadotropins (FSH and/or LH) from the anterior pituitary.
Cows also have periods of anestrus (noncycling). For example, an anestrous period is commonly observed in cows following calving. Low levels of nutrition can contribute to the duration of anestrus, especially in young cows nursing calves.
Estrus is not always accompanied by ovulation, nor ovulation by estrus. Heat without ovulation (anovulatory heat) will not result in pregnancy, even if the female is bred. Ovulation without the external signs of heat (quiet or silent heats) is not uncommon in cows, especially the first few weeks after calving. Such females will generally not "stand" to be bred by a bull.
The discussion of the hormonal control of the estrous cycle and pregnancy as presented here is a greatly simplified summary of the whole process. In reality, the reproductive process is very complicated and involves a number of hormonal interactions and events.
References
Deutscher, G.H. 1980. Reproductive tract anatomy and physiology of the cow [G80-537-A]. Lincoln: University of Nebraska Cooperative Extension.
Rich, T.D., and E.J. Turman. n.d. Reproductive tract anatomy and physiology of the cow [Beef Cattle Handbook BCH-2200]. Stillwater: Oklahoma State University.
Original authors: Ron Parker and Clay Mathis, Extension Livestock Specialists. Adapted from Extension Beef Cattle Resource Committee Beef Cattle Handbook publication BCH-2200, by T.D. Rich and E.J. Turman, Animal Science Department, Oklahoma State University.

Jason L. Turner is Associate Professor and Extension Horse Specialist. Jason was active in 4-H and FFA while growing up in Northeastern Oklahoma. His M.S. and Ph.D. studies concentrated on equine reproduction, health, and management. His Extension programs focus on proper care and management of the horse for youth and adults.
To find more resources for your business, home, or family, visit the College of Agricultural, Consumer and Environmental Sciences on the World Wide Web at pubs.nmsu.edu
Contents of publications may be freely reproduced for educational purposes. All other rights reserved. For permission to use publications for other purposes, contact pubs@nmsu.edu or the authors listed on the publication.
New Mexico State University is an equal opportunity/affirmative action employer and educator. NMSU and the U.S. Department of Agriculture cooperating.
Revised July 2014 Las Cruces, NM